X




|
STUDENT DIGITAL NEWSLETTER ALAGAPPA INSTITUTIONS |

|
Jonathan D. Kirsch, MD
Various abnormal haemoglobins (Hb variants) are due to changes in the genetic code treatment writing discount detrol 2 mg amex. Some codons do not code for any amino acid and they cause termination of polypeptide chain formation medicine recall discount 1mg detrol otc. Existence of more than one code word for an amino acid is known as degeneracy of genetic code symptoms before period discount detrol 2 mg online. Further medications ending in pril order detrol 1mg line, codon and anticodons are anti-parallel and complementary in base composition acne natural treatment buy cheap detrol 4mg. For proper function of codon symptoms anemia cheap 2 mg detrol free shipping, only first two nucleotides are essential the third nucleotide is flexible, i. Thus, the base pairing of third nucleotide of codon with anti-codon is less specific. They are called as ribonucleo proteins because they are rich in ribonucleic acids. Structure of ribosomes Like proteins ribosomes have characteristic three-dimensional structure. Mechanism of protein biosynthesis Rough endoplasmic reticulum is the site of formation of proteins in eukaryotes and cytosol is the site of protein synthesis in prokaryotes. Like transcription, mechanism of translation involves initiation, elongation and termination. Each of three processes require several proteins, which are different in prokaryotes and eukaryotes. Coli and in all other prokaryotes starting amino acid is always N-formyl methionine. Activation of amino acids For protein synthesis, initial activation of amino acid is required. This activation is essential because energy is required for peptide bond formation. In the second reaction a formyl group is transferred to the methionine from N10-formyl tetra hydrofolate by specific transformylase enzyme (Figure 18. Elongation Now the polypeptide chain is elongated by covalent attachment of successive amino acids from amino terminus to carboxy terminus. The peptidyl transferase activity of 50S ribosomal subunit catalyzes the peptide bond formation between two amino acids. The elongation process is repeated several times adding one amino acid each time until a stop codon is encountered in the A site. Chloramphenicol It inhibits protein biosynthesis by blocking peptidyl transferase activity of 50S subunit. Tunicamycin It prevents attachments of oligosaccharide side chains to certain glyco proteins. Cyclo heximide It inhibits protein biosynthesis in eukaryotes by blocking peptidyl transferase activity of 60S eukaryotic ribosome. Translational factors as chaperones Several translational factors also possess chaperone activity. They are ancestral chaperones dedicated to folding of nascent polypeptides before the evolution of general chaperones. Protein targeting Now we shall examine how proteins are transported to various locations in the cell from the site of synthesis where they function as cytosolic enzyme, membrane components, digestive enzymes, hormones etc. Proteins have intrinsic signals or molecular addresses that directs their transport and localization in the cell. Secretary proteins are translated on membrane bound ribosomes where as cytosolic proteins are synthesized on free ribosomes. Translation of signal sequence results in unique amino acid sequence on amino terminal end of nascent polypeptide. These signals has all information for targeting proteins to organelles, translocation across membrane or assembly into membranes. Protein sorting pathway of secretary proteins begins with synthesis of signal peptide by free ribosomes in cytosol. A ribosome receptor, translocon of endoplasmic reticulum aids passage of peptide chain through lipid bilayer and synthesis proceeds on membrane-bound ribosomes. Usually protein may attain proper conformation by the time it is released from the ribsome. Phosphorylation the hydroxyl groups of serine and tyrosine residues of some proteins are phosphorylated after they leave ribosome. For example, casein has many phosphate groups, which are attached to hydroxyl group of serine after it comes out of ribosomes. For example, phosphorylation of tyrosine residues of certain proteins converts normal cells to cancer cells. Attachment of prosthetic groups Initially protein part of conjugated protein is synthesized on ribosome. Hydroxylation Proline and lysine residues of collagen subunits are hydroxylated after they leaves ribosomes. Disulfide bond formation In some proteins, inter or intrachain disulfide bonds are formed after proteins leaves ribosome. Methylation In some proteins, lysine residues are methylated after they leaves ribosomes. Carboxylation Carboxyl groups are added to glutamic and aspartic acid residues of some proteins after they leaves ribosome. For example, -carboxylation of glutamyl residues of blood clotting factors like prothrombin. Removal of initiating amino acid the initiating amino acid is removed in some proteins as soon as they comes out of ribosome. Proteolytic modifications Some proteins that are going to be exported from the cell undergo proteolytic modification. Palmitoylation It is the covalent attachment of lipid moieties to proteins that is found in prokaryotes and eukaryotes. Palmitic acid is the lipid post translationally attached to intergral and peripheral membrane proteins. N-Palmitoylation In which palmitate is attached to amino terminal cysteine of the protein through amide linkage. Functions of palmitoylation Functions of palmitoylation depends on protein that is being considered. For example targeting of proteins to axons and dendrites in neurons is influenced by palmitoylation. Palmitoylation facilitates transport of newly synthesized chemokine receptor to plasma membrane. For example, palmitoylation is crucial for binding of erythrocyte membrane protein 4. Methyl malonyl semi aldehyde dehydrogenase is another enzme whose activity is regulated by palmitoylation. Medical importance Mutations in genes results in nonfunctional protein production and cancer development. They may be Transitions In which a purine base is changed to another purine base or a pyrimidine base is changed to another pyrimidine bases. Some times normal protein may be produced because of the degeneracy of genetic code. Missense mutation In which a wrong amino acid is incorporated into protein due to changed base in codon. Wrong amino acid in protein may not affect function of protein, can partially affect function of protein, abolish protein function. Acceptable missense mutation Haemoglobin Hikari is an example for acceptable missense mutation. As a result in 61 position of chain aspargine is incorporated in place of lysine and abnormal haemoglobin Hb Hikari is produced. Partially acceptable missense mutation HbS is an example for partially acceptable missense mutation. Unacceptable missense mutation HbM (Boston) is an example for this type of mutation. Nonsense mutation In which a normal codon is converted to termination (nonsense) codon due to change of base in codon. Deletion mutation Abnormal haemoglobin, Hb Wayne is an example for deletion mutation. As a result, the chain is elongated beyond its normal length and the -chain of Hb Wayne contains 146 amino acids instead of normal 141 residues. Insertion mutation Abnormal haemoglobin Hb Cranston is an example for insertion mutation. As a result, chain is elongated beyond its normal length and chain of Hb Cranston contains 157 amino acids instead of normal 146 amino acids. Oncogene identified for bladder carcinoma differs from normal gene in only one nucleotide substitution. Isolation and Identification of short nucleotide sequences that affects translation initiation in S. Diphtheria toxin inhibits protein synthesis (a) By inactivating initiation factor. Gene expression is subjected to regulation throughout the life span of prokaryotic and eukaryotic cells. Expression of genes is turned on or turned off depending on biological needs, developmental stage or time, environmental (external) and nutritional factors. For example, when cell prepares for division only genes that have role in cell division are expressed like genes which code replication enzymes and genes that code proteins which are essential for proper cell division. After cell divides expression of these genes is turned off and expression of genes that are involved in various house keeping activities or maintenance of cell is turned on. Further, the extent of expression of genes that are involved in various cellular processes also differs. When cell completes its life span apoptosis occurs due to expression of certain genes associated with it. Development of human embryo to infant involves turning on or tuning off of several genes. Growth of infant into adult involves excessive expression of genes that are essential for growth. Even though all eukaryotic cells contain same information, its expression depends on tissue. Depending on environmental or nutritional factors, expression of certain genes is increased or decreased. For example, expression of key enzymes of gluconeogenesis is increased in starvation like wise when toxic substances are consumed expression of certain genes increases to destroy these toxic substances. In contrast when a particular substance is present in cellular environment at definite concentration 459 460 Medical Biochemistry expression of certain genes is diminished. When bacteria is present in a medium lacking specific amino acid expression of certain genes increases. In contrast when bacteria is present in a medium rich in specific amino acid expression of genes, which are involved in its synthesis decreases. Proper understanding of hormonal regulation of gene expression is essential for development of drugs or treatment. Antisense molecules can block gene expression, which is the basis for antisense therapy. Maintenance of proper intracellular metal level involves turning on or off of gene expression. We shall first examine gene expression in prokaryotes then proceed to eukaryotic gene regulation. However, gene regulation in eukaryotes at molecular level is not known completely in several processes. Increased expression of genes results in increased concentration of enzymes because enzymes are gene products. Under normal conditions, these enzymes are present in small amounts but their concentration raises to many folds in presence of inducer due to increased expression of genes. Repression Diminished expression of genes in presence of repressor is called as repression. Lac operon Model To explain gene regulation by induction and repression, Jacob and Monod proposed operon model. This model explains regulation of gene expression at transcriptional and translational levels. These structural genes codes for - galactosidase, permease and transacetylase respectively. As a result the enzymes involved in lactose, catabolism are not formed or repressed. When present lactose (inducer) binds to repressor molecule and forms inducer-repressor complex. So, formation of inducer-repressor complex prevents repressor to bind with operator gene. Coli is grown in medium containing glucose and lactose, lac operon is turned off and cells use exclusively glucose. The inhibitory action of glucose on lac operon is called as catabolite repression. Now we shall examine an operon related to biosynthesis to know whether it is similar to lac operon. Since biosynthesis consumes energy usually biosynthesis operon is turned off when compound is present in medium. We shall examine biosynthesis operon related to synthesis of tryptophan to know how tryptophan in medium can turn off its own synthesis. Since the operon is related to tryptophan metabolism it is called as tryptophan operon or try operon. When tryptophan is present in medium it acts as corepressor and binds to aporepressor to form holo repressor, which binds to operator gene.
Some plasmids promote mating between bacteria; others contain genes that kill other bacteria symptoms jaw bone cancer order detrol 4 mg with mastercard. Of great importance symptoms zika virus order 2 mg detrol with mastercard, plasmids are used extensively in genetic engineering (see Chapter 19) medications covered by medi cal discount detrol 1mg on-line, and some of them play a role in the spread of antibiotic resistance among bacteria symptoms 7 dpo bfp cheap detrol 4 mg overnight delivery. The origin allows a plasmid to replicate independently of the bacterial chromosome (Figure 8 medications side effects discount detrol 1mg line. Episomes are plasmids that are capable of freely replicating and able to integrate into the bacterial chromosomes medicine express order detrol 4mg mastercard. A typical bacterial genome consists of a single circular chromosome that contains several million base pairs. Replication begins at the origin of replication (ori) and continues around the circle. In this diagram, replication is taking place in both directions; in some plasmids, replication is in one direction only. Most bacteriophages have a limited host range; so transduction is normally between bacteria of the same or closely related species only. These processes of genetic exchange in bacteria differ from diploid eukaryotic sexual reproduction in two important ways. Each type of genetic transfer can be used to map genes, as will be discussed in the following sections. Each type of genetic transfer consists of a one-way movement of genetic information to the recipient cell, sometimes followed by recombination. Conjugation takes place when genetic material passes directly from one bacterium to another (Figure 8. In conjugation, two bacteria lie close together and a connection forms between them. A plasmid or a part of the bacterial chromosome passes from one cell (the donor) to the other (the recipient). After transformation, recombination may take place between the introduced genes and those of the bacterial chromosome. All of the above Conjugation In 1946, Joshua Lederberg and Edward Tatum demonstrated that bacteria can transfer and recombine genetic information, paving the way for the use of bacteria in genetic studies. In the course of their research, Lederberg and Tatum studied auxotrophic strains of E. The Y10 strain required the amino acids threonine (and was genotypically thr-) and leucine (leu-) and the vitamin thiamine (thi-) for growth but did not require the vitamin biotin (bio+) or the amino acids phenylalanine (phe+) and cysteine (cys+); the genotype of this strain can be written as thr- leu- thi- bio+ phe+ cys+. The Y24 strain had the opposite set of alleles: it required biotin, phenylalanine, and cysteine in its medium, but it did not require threonine, leucine, or thiamine; its genotype was thr+ leu+ thi+ bio- phe- cys-. In one experiment, Lederberg and Tatum mixed Y10 and Y24 (a) Conjugation Donor cell Recipient cell Cytoplasmic bridge forms. Strain Y10 was unable to grow, because it required threonine, leucine, and thiamine, which were absent in the minimal medium; strain Y24 was unable to grow, because it required biotin, phenylalanine, and cysteine, which also were absent from the minimal medium. When Lederberg and Tatum mixed the two strains, however, a few colonies did grow on the minimal medium. If mutations were responsible for the prototrophic colonies, then some colonies should also have grown on the plates containing Y10 or Y24 alone, but no bacteria grew on these plates. Multiple simultaneous mutations (thr- thr+, leu- leu+, and thi- thi+ in strain Y10 or bio- bio+, phe- phe+, and cys- cys+ in strain Y24) would have been required for either strain to become prototrophic by mutation, which was very improbable. Lederberg and Tatum concluded that some type of genetic transfer and recombination had taken place: 209 210 Chapter 8 Experiment Question: Do bacteria exchange genetic information Strain A Strain B Results + + leu + thi bio phe + thr + cys + Two auxotrophic strains were separated by a filter that allowed mixing of medium but not bacteria. Results Minimal medium Minimal medium Minimal medium Minimal medium Conclusion: Yes, genetic exchange and recombination took place between the two mutant strains. No growth No growth No growth No growth Conclusion: Genetic exchange requires direct contact between bacterial cells. This filter allowed liquid medium to pass from one side of the tube to the other, but the pores of the filter were too small to allow the passage of bacteria. Two auxotrophic strains of bacteria were placed on opposite sides of the filter, and suction was applied alternately to the ends of the U-tube, causing the medium to flow back and forth between the two compartments. Despite hours of incubation in the U-tube, bacteria plated out on minimal medium did not grow; there had been no genetic exchange between the strains. The exchange of bacterial genes clearly required direct contact, or conjugation, between the bacterial cells. The F factor contains an origin of replication and a number of genes required for conjugation (see Figure 8. For example, some of these genes encode sex pili (singular, pilus), slender extensions of the cell membrane. A cell containing F produces the sex pili, one of which makes contact with a receptor on an F- cell (Figure 8. Conjugation can take place only between a cell that possesses F and a cell that lacks F. In most cases, the only genes transferred during conjugation between an F+ and F- cell are those on the F factor (Figure 8. Replication takes place on the nicked strand, proceeding around the circular plasmid in the F+ cell and replacing the transferred strand (Figure 8. Because the plasmid in the F+ cell is always nicked at the oriT site, this site always enters the recipient cell first, followed by the rest of the plasmid. Inside the recipient cell, the single strand replicates, producing a circular, double-stranded copy of the F plasmid (Figure 8. If the entire F factor is transferred to the recipient F- cell, that cell becomes an F+ cell. F+ and F- cells In most bacteria, conjugation depends on a fertility (F) factor that is present in the donor cell and Hfr cells Conjugation transfers genetic material in the F plasmid from F+ to F- cells but does not account for the transfer of chromosomal genes observed by Lederberg and Tatum. In Hfr (high-frequency) strains, the F factor is integrated into the bacterial chromosome (Figure 8. Hfr cells behave as F+ cells, forming sex pili and undergoing conjugation with F- cells. Because, in an Hfr cell, the F factor has been integrated into the bacterial chromosome, the chromosome follows it into the recipient cell. How much of the bacterial chromosome is transferred depends on the length of time that the two cells remain in conjugation. This gene transfer between Hfr and F- cells is how the recombinant prototrophic cells observed by Lederberg and Tatum were produced. After crossing over has taken place in the recipient cell, the donated chromosome is degraded and the recombinant recipient chromosome remains (Figure 8. To become F+ or Hfr, the recipient cell must receive the entire F factor, requiring the entire bacterial chromosome to be transferred. This event happens rarely, because most conjugating cells break apart before the entire chromosome has been transferred. The F plasmid in F+ cells integrates into the bacterial chromosome, causing an F+ cell to become Hfr, at a frequency of only about 1/10,000. This low frequency accounts for the low rate of recombination observed by Lederberg and Tatum in their F+ cells. The F factor is excised from the bacterial chromosome at a similarly low rate, causing a few Hfr cells to become F+. F cells When an F factor does excise from the bacterial chromosome, a small amount of the bacterial chromosome may be removed with it, and these chromosomal genes will then be carried with the F plasmid (Figure 8. For example, if an F factor integrates into a chromosome adjacent to the lac genes (genes that enable a cell to metabolize the sugar lactose), the F factor may pick up lac genes when it excises, becoming Flac. Crossing over may lead to the recombination of alleles (bright green in place of black segment). Bacterial and Viral Genetic Systems 213 Crossing over takes place within the Hfr chromosome. During conjugation between an Flac cell and an F- cell, the F plasmid is transferred to the F- cell, which means that any genes on the F plasmid, including those from the bacterial chromosome, may be transferred to F- recipient cells. It produces partial diploids, or merozygotes, which are cells with two copies of some genes, one on the bacterial chromosome and one on the newly introduced F plasmid. Cells containing F (F+ cells) are donors during gene transfer; cells lacking F (F- cells) are recipients. Chromosome transfer always begins within the integrated F factor and proceeds in a continuous direction; so genes are transferred according to their sequence on the chromosome. The time required for individual genes to be transferred indicates their relative positions on the chromosome. In most genetic maps, distances are expressed as percent recombination; but, in bacterial maps constructed with interrupted conjugation, the basic unit of distance is a minute. They used donor Hfr cells that were sensitive to the antibiotic streptomycin (genotype str s), resistant to sodium azide (azir) and infection by bacteriophage T1 (tonr), prototrophic for threonine (thr+) and leucine (leu+), and able to break down lactose (lac+) and galactose (gal+). They used F- recipient cells that were resistant to streptomycin (str r), sensitive to sodium azide (azi s) and to infection by bacteriophage T1 (tons), auxotrophic for threonine (thr-) and leucine (leu-), and unable to break down lactose (lac-) and galactose (gal-). Thus, the genotypes of the donor and recipient cells were: Donor Hfr cells: str leu thr azi ton lac gal - r - - s s - s + + r r + + - Question: How can interrupted conjugation be used to map bacterial genes Genes transferred: str s, leu +, and thr + (first selected genes, defined as zero time) 3 Conjugation was interrupted at regular intervals. The cells from each sample were plated on a selective medium that contained streptomycin and lacked leucine and threonine. The original donor cells were streptomycin sensitive (str s) and would not grow on this medium. The F- recipient cells were auxotrophic for leucine and threonine, and they also failed to grow on this medium. Only cells that underwent conjugation and received at least the leu+ and thr+ genes from the Hfr donors could grow on this medium. All str r leu+ thr+ cells were then tested for the presence of other genes that might have been transferred from the donor Hfr strain. All of the cells that grew on this selective medium were str r + leu thr+; so we know that these genes were transferred. The percentage of str r leu+ thr+ cells receiving specific alleles (azir, tonr, leu+, and gal +) from the Hfr strain are plotted against the duration of conjugation (Figure 8. What are the order in which the genes are transferred and the distances among them Gene ton r appeared next (after about 10 minutes), followed by lac+ (at about 18 minutes) and by gal + (after 25 minutes). These transfer times indicate the order of gene transfer and the relative distances among the genes (see Figure 8. For example, about 90% of the recipients received the azir allele, but only about 30% received the gal+ allele. The lower percentage for gal+ is due to the fact that some conjugating cells spontaneously broke apart before they were disrupted by the blender. The probability of spontaneous disruption increases with time; so fewer cells had an opportunity to receive genes that were transferred later. For additional practice mapping bacterial genes with interrupted conjugation, try Problem 21 at the end of the chapter. Gene transfer always begins within F, and the orientation and position of F determine the direction and starting point of gene transfer. Genes from this strain will be transferred in the order of: leu-thr-thi-his-gal-lac-pro-azi In strain Hfr5, F is integrated between the thi and the his genes (Figure 8. Here gene transfer will proceed in a clockwise direction: thi-thr-leu-azi-pro-lac-gal-his Although the starting point and direction of transfer may differ between two strains, the relative distance in time between any two pairs of genes is constant. Notice that the order of gene transfer is not the same for different Hfr strains (Figure 8. For example, azi is transferred just after leu in strain HfrH but long after leu in strain Hfr1. That they are the same makes sense when we recognize that the bacterial chromosome is circular and the starting point of transfer varies from strain to strain. These data provided the first evidence that the bacterial chromosome is circular (Figure 8. The donor genes first appeared in the recipient cells at the following times: gal, 10 minutes; his, 8 minutes; pro, 15 minutes. The amount of time required for individual genes - to be transferred from the Hfr to the F cells indicates the relative positions of the genes on the bacterial chromosome. Natural Gene Transfer and Antibiotic Resistance Antibiotics are substances that kill bacteria. Their development and widespread use has greatly reduced the threat of infectious disease and saved countless lives. But many pathogenic bacteria have developed resistance to antibiotics, 216 Chapter 8 (a) Hfr strain thr leu azi pro lac gal his H leu thr thi 1 pro azi leu thr thi 2 lac pro azi leu thr thi 3 thi 4 thi thr leu azi pro lac gal his 5 (c) thr thi his gal lac 5 leu azi pro thi his gal lac H thr leu azi pro thi his gal his gal lac pro azi leu thr his gal his gal lac his gal lac pro azi thi (b) Hfr strain thi thr leu azi pro lac gal his 5 thr leu azi pro lac gal his H thr leu azi pro lac gal his 4 azi pro lac gal his 1 lac gal his 2 gal his 3 thi thr leu azi pro lac thi thr leu azi pro thi thr leu thi thi thr leu 4 azi pro lac thi his gal thr leu 1 azi pro lac thi his gal thr leu 2 azi pro lac thi his gal thr leu 3 azi pro lac Conclusion: the order of the genes on the chromosome is the same, but the position and orientation of the F factor differ among the strains. In these enviornments where antibiotics are continually present, the only bacteria to survive are those that possess resistance to antibiotics. No longer in competition with other bacteria, resistant bacteria multiply quickly and spead. In this way, the presence of antibiotics selects for resistant bacteria and reduces the effectiveness of antibiotic treatment for medically important infections.
The original copy can provide the essential function medications qhs detrol 2 mg amex, whereas an extra copy from the duplication is free to undergo mutation and change symptoms 8dp5dt cheap detrol 4 mg with visa. Over evolutionary time medications not covered by medicare trusted 1mg detrol, the extra copy may acquire enough mutations to assume a new function that benefits the organism treatment genital herpes discount 2mg detrol with amex. Inversions also can play important evolutionary roles by suppressing recombination among a set of genes medications bad for your liver buy generic detrol 4mg on-line. As we have seen symptoms you need a root canal generic detrol 2 mg on-line, crossing over within an inversion in an individual Source: After F. Problems in chromosome pairing and segregation often lead to sterility in autopolyploids, but many allopolyploids are fertile. This suppression of recombination allows particular sets of coadapted alleles that function well together to remain intact, unshuffled by recombination. Polyploidy, particularly allopolyploidy, often gives rise to new species and has been particularly important in the evolution of flowering plants. Occasional genome doubling through polyploidy has been a major contributor to evolutionary suc- cess in animal groups. For example, Saccharomyces cerevisiae (yeast) is a tetraploid, having undergone whole-genome duplication about 100 million years ago. The vertebrate genome has duplicated twice, once in the common ancestor to jawed vertebrates and again in the ancestor of fishes. Certain groups of vertebrates, such as some frogs and some fishes, have undergone additional polyploidy. Duplications often have pronounced effects on the phenotype owing to unbalanced gene dosage. In individuals heterozygous for an inversion, the homologous chromosomes form inversion loops in meiosis, with reduced recombination taking place within the inverted region. These variants are common in the human genome; some are associated with diseases and disorders. Mosaicism is caused by nondisjunction in an early mitotic division that leads to different chromosome constitutions in different cells of a single individual. It is produced when the dominant wild-type allele in a heterozygous individual is absent due to a deletion on one chromosome. Dosage compensation prevents the expression of additional copies of X-linked genes in mammals, and there is little information in the Y chromosome; so extra copies of the X and Y chromosomes do not have major effects on development. In contrast, there is no mechanism of dosage compensation for autosomes, and so extra copies of autosomal genes are expressed, upsetting development and causing the spontaneous abortion of aneuploid embryos. Give the chromosomes that would be found in individuals with the following chromosome mutations. Because the centromere has not changed location and is not in the inverted region, this chromosome mutation is a paracentric inversion. Sketch the chromosome pairing and the different segregation patterns that can arise in an individual heterozygous for a reciprocal translocation. Draw a pair of chromosomes as they would appear during synapsis in prophase I of meiosis in an individual heterozygous for a chromosome duplication. Explain, with the aid of a drawing, how a dicentric bridge is produced when crossing over takes place in an individual heterozygous for a paracentric inversion. Explain why recombination is suppressed in individuals heterozygous for paracentric and pericentric inversions. Explain why autopolyploids are usually sterile, whereas allopolyploids are often fertile. For each mutation, sketch how the wild-type and mutated chromosomes would pair in prophase I of meiosis, showing all chromosome strands. Draw the chromatids that would result from a two-strand double crossover between E and F in Problem 24. Maarit Jaarola and colleagues examined individual sperm cells of a male who was heterozygous for a pericentric inversion on chromosome 8 and determined that crossing over took place within the pericentric inversion in 26% of the meiotic divisions (M. Assume that you are a genetic counselor and that a couple seeks counseling from you. Both the man and the woman are phenotypically normal, but the woman is heterozygous for a pericentric inversion on chromosome 8. What is the probability that this couple will produce a child with a debilitating syndrome as the result of crossing over within the pericentric inversion Diagram the alternate, adjacent-1, and adjacent-2 segregation patterns in anaphase I of meiosis. Give the products that result from alternate, adjacent-1, and adjacent-2 segregation. Female flies heterozygous for Notch have an indentation on the margins of their wings; Notch is lethal in the homozygous and hemizygous conditions. The Notch deletion covers the region of the X chromosome that contains the locus for white eyes, an X-linked recessive trait. The green-nose fly normally has six chromosomes, two metacentric and four acrocentric. A geneticist examines the chromosomes of an odd-looking green-nose fly and discovers that it has only five chromosomes; three of them are metacentric and two are acrocentric. Where did the nondisjunction that gave rise to the young man with Klinefelter syndrome take place Assuming that no new mutations occurred in this family, explain the presence of an autosomal recessive disease in the child when the mother is heterozygous and the father is homozygous normal. On the basis of these observations, indicate which of the following statements are most likely correct and which are most likely incorrect. In mammals, sex-chromosome aneuploids are more common than autosomal aneuploids but, in fish, sexchromosome aneuploids and autosomal aneuploids are found with equal frequency. A chromosome analysis reveals that, whereas the woman has a normal karyotype, the a. What types of zygotes will develop when each of gametes produced by the man fuses with a normal gamete produced by the woman If trisomies and monosomies entailing chromosomes 13 and 22 are lethal, what proportion of the surviving offspring will be carriers of the translocation Oxford: Oxford University Press, Clarendon Press; New York: Oxford University Press, 1987) created mice that were trisomic for each of the different mouse chromosomes. For some of these trisomics, they compared the length of development (number of days after conception before the embryo died) as a function of the size of the mouse chromosome that was present in three copies (see the adjoining graph). Summarize their findings as presented in this graph and provide a possible explanation for the results. How many chromosomes will be found per cell in each of the following mutants in this species What types of chromosome mutations do individual organisms with the following sets of chromosomes have What would the expected chromosome numbers be in individual organisms with the following chromosome mutations Consider a diploid cell that has 2n = 4 chromosomes- one pair of metacentric chromosomes and one pair of acrocentric chromosomes. Suppose this cell undergoes nondisjunction giving rise to an autotriploid cell (3n). Draw the different types of gametes that may result from meiosis in the triploid cell, showing the chromosomes present in each type. To distinguish between the different metacentric and acrocentric chromosomes, use a different color to draw each metacentric chromosome; similarly, use a different color to draw each acrocentric chromosome. They were able to self-pollinate the flowers of this plant to produce an F2 generation. What would be the chromosome number of progeny resulting from the following crosses in wheat (see Figure 9. Einkorn wheat and emmer wheat Bread wheat and emmer wheat Einkorn wheat and bread wheat Karl and Hally Sax crossed Aegilops cylindrica (2n = 28), a wild grass found in the Mediterranean region, with Triticum vulgare (2n = 42), a type of wheat (K. Examination of metaphase I in the F1 plants revealed the presence of 7 pairs of chromosomes (bivalents) and 21 unpaired chromosomes (univalents). If the unpaired chromosomes segregate randomly, what possible chromosome numbers will appear in the gametes of the F1 plants What does the appearance of the bivalents in the F1 hybrids suggest about the origin of Triticum vulgare wheat James and Margaret Lesley observed that secondaries arise from triploid (3n), trisomic (3n + 1), and double trisomic (3n + 1 + 1) parents, but never from diploids (2n). Give one or more possible reasons that secondaries arise from parents that have unpaired chromosomes but not from parents that are normal diploids. Blood tests established that the male foal, appropriately named Blue Moon, was the offspring of Krause and that Krause was indeed a mule. Both Blue Moon and Krause were fathered by the same donkey (see the illustration). The foal, like his mother, had 63 chromosomes-half of them horse chromosomes and the other half donkey chromosomes. Analyses of genetic markers showed that, remarkably, Blue Moon seemed to have inherited a complete set of horse chromosomes from his mother, instead of the random mixture of horse and donkey chromosomes that would be expected with normal meiosis. Thus, Blue Moon and Krause were not only mother and son, but also brother and sister. With the use of a diagram, show how, if Blue Moon inherited only horse chromosomes from his mother, Blue Moon and Krause are both mother and son as well as brother and sister. Although rare, additional cases of fertile mules giving births to offspring have been reported. In these cases, when a female mule mates with a male horse, the offspring is horselike in appearance but, when a female mule mates with a male donkey, the offspring is mulelike in appearance. Is this observation consistent with the idea that the offspring of fertile female mules inherit only a set of horse chromosomes from their mule mothers Can you suggest a possible mechanism for how the offspring of fertile female mules might pass on a complete set of horse chromosomes to their offspring Humans and many other complex organisms are diploid, possessing two sets of genes, one inherited from the mother and one from the father. However, a number of eukaryotic organisms spend most of their life cycles in a haploid state. Many of these eukaryotes, such as Neurospora and yeast, still undergo meiosis and sexual reproduction, but most of the cells that make up the organism are haploid. Considering that haploid organisms are fully capable of sexual reproduction and generating genetic variation, why are most complex eukaryotes diploid In other words, what might be the evolutionary advantage of existing in a diploid state instead of a haploid state In 1856, a group of men working a limestone quarry in the Neander Valley of Germany discovered a small cave containing a number of bones. The workers assumed that the bones were those of a cave bear, but a local schoolteacher recognized them as human, although they were clearly unlike any human bones the teacher had ever seen. The bones appeared to be those of a large person with great muscular strength, a low forehead, a large nose with broad nostrils, and massive protruding brows. Experts confirmed that the bones belonged to an extinct human, who became known as Neanderthal. In the next 100 years, similar fossils were discovered in Spain, Belgium, France, Croatia, and the Middle East. Research has now revealed that Neanderthals roamed Europe and western Asia for at least 200,000 years, disappearing abruptly 30,000 to 40,000 years ago. During the last years of this period, Neanderthals coexisted with the direct ancestors of modern humans, the Cro-Magnons. The fate of the Neanderthals-why they disappeared-has captured the imagination of scientists and laypersons alike. Did Cro-Magnons, migrating out of Africa with a superior technology, cause the demise of the Neanderthal people, either through competition or perhaps through deliberate extermination Or did the Neanderthals interbreed with Cro-Magnons, their genes becoming assimilated into the larger gene pool of modern humans Support for the latter hypothesis came from the discovery of fossils that appeared to be transitional between Neanderthals and Cro-Magnons. Unfortunately, the meager fossil record of Neanderthals and Cro-Magnons did not allow a definitive resolution of these questions. Comparison of the genomes of modern humans and Neanderthals reveal evidence of some interbreeding between the two groups.
Where more than one cross was completed for a pair of genes treatment tinea versicolor cheap 4mg detrol free shipping, average the recombination rates from the different crosses medications not to mix detrol 1mg generic. Two mutations that affect plaque morphology in phages (a- and b-) have been isolated treatment yellow jacket sting order detrol 2 mg free shipping. Phages carrying both mutations (a- b-) are mixed with wild-type phages (a+ b+) and added to a culture of bacterial cells medications to treat bipolar disorder order detrol 4 mg line. Subsequent to infection and lysis medicine man dispensary cheap detrol 1 mg online, samples of the phage lysate are collected and cultured on bacterial cells medications quit smoking generic 1mg detrol amex. The following numbers of plaques are observed: Plaque phenotype a b a+ b- a- b+ a- b- + + On the basis of these data, which gene is farthest from his2+ Seven of the strains that they used in their study are listed here, along with the mutation found in that strain. On the basis of complementation studies, they found four proline auxotrophs: proA, proB, proC, and proD. To determine whether proA, proB, proC, and proD loci were located close together on the bacterial chromosome, they conducted a transduction experiment. Bacterial strains that were proC + and had mutations at proA, proB, or proD, were used as donors. The bacteria were then plated on a selective medium that allowed only proC + bacteria to grow. The following results were obtained: Donor genotype Transductant genotype Number 2765 3 1838 2 1166 0 To map the genes for tryptophan synthesis, they carried out a series of transformation experiments on strains having different mutations and determined the percentage of recombinants among the transformed bacteria. Which genotypes represent single transductants and which represent cotransductants Previous mapping experiments have established that the genes responsible for these two mutations are 8 m. The geneticist mixes phages with genotype c+ m+ and genotype c- m- and uses the mixture to infect bacterial cells. What numbers of the different types of plaques (c+ m+, c- m-, c+ m-, c- m+) should she expect to see The geneticist carries out the same experiment described in Problem 30, but this time she mixes phages with genotypes c+ m- and c- m+. A geneticist isolates two bacteriophage r mutants (r13 and r2) that cause rapid lysis. He carries out the following crosses and counts the number of plaques listed here: Genotype of parental phage h + Progeny phage genotype h c h c h+ c h c+ h+ c h c+ h+ c+ h c + + Number of plaques 321 338 26 30 106 110 5 6 st st st st+ st+ st st st+ + a. A donor strain of bacteria with genes a+ b+ c+ is infected with phages to map the donor chromosome with generalized transduction. The phage lysate from the bacterial cells is collected and used to infect a second strain of bacteria that are a- b- c-. Bacteria with the a+ gene are selected, and the percentage of cells with cotransduced b+ and c+ genes are recorded. The phage lysate from the bacterial cells is collected and used to infect a second strain of bacteria that are leu- gal+ pro-. One strain has a mutant host range, is temperature sensitive, and produces clear plaques (genotype is h st c); another strain carries the wild-type alleles (genotype is h+ st+ c+). The genotypes of the progeny phages are given here: Donor leu gal pro + - + Recipient leu- gal+ pro- Which genes are closest, leu and gal or leu and pro To determine whether these mutations occur at the same functional gene, he simultaneously infects E. As a summer project, a microbiology student independently isolates two mutations in E. The student wants to know whether these two mutants are at the same functional unit. Outline a procedure that the student could use to determine whether these two gly- mutations occur within the same functional unit. A group of genetics students mix two auxotrophic strains of bacteria: one is leu+ trp+ his- met- and the other is leu- trp- his+ met+. After mixing the two strains, they plate the bacteria on minimal medium and observe a few prototrophic colonies (leu+ trp+ his+ met+). How can they determine whether the transfer of genes is due to conjugation, transduction, or transformation Their features were so similar, in fact, that he felt that they might easily be mistaken as children of the same family. Down did not understand the cause of their retardation, but his original description faithfully records the physical characteristics of this most common genetic form of mental retardation. As early as the 1930s, geneticists suggested that Down syndrome might be due to a chromosome abnormality, but not until 1959 did researchers firmly establish the cause of Down syndrome: most people with the disorder have three copies of chromosome 21, a condition known as trisomy 21. In a few rare cases, people having the disorder are trisomic for just specific parts of chromosome 21. In spite of this exciting finding, the genetics of Down syndrome appears to be more complex than formerly thought. Mouse breeders have developed several strains of mice that are trisomic for most of the genes found on human chromosome 21 (the equivalent mouse genes are found on mouse chromosome 16). These mice display many of the same anatomical features found in people with Down syndrome, as well as altered behavior, and they are considered an animal model for Down syndrome. This gene appears to be responsible for at least some of the Alzheimer-like features observed in older Downsyndrome people. Taken together, findings from these studies suggest that Down syndrome is not due to a single gene but is instead caused by complex interactions among multiple genes that are affected when an extra copy of chromosome 21 is present. Research on Down syndrome illustrates the principle that chromosome abnormalities often affect many genes that interact in complex ways. Nevertheless, variations in chromosome number-such as the extra chromosome 21 that leads to Down syndrome-do periodically arise. Variations may also arise in chromosome structure: individual chromosomes may lose or gain parts and the order of genes within a chromosome may become altered. These variations in the number and structure of chromosomes are termed chromosome mutations, and they frequently play an important role in evolution. We begin this chapter by briefly reviewing some basic concepts of chromosome structure, which we learned in Chapter 2. We then consider the different types of chromosome mutations, their definitions, features, phenotypic effects, and influence on evolution. M Chromosome Morphology Each functional chromosome has a centromere, to which spindle fibers attach, and two telomeres, which stabilize the chromosome (see Figure 2. The centromere is located approximately in the middle, and so the chromosome has two arms of equal length. The centromere is near one end, producing a long arm and a knob, or satellite, at the other end. The complete set of chromosomes possessed by an organism is called its karyotype and is usually presented as a picture of metaphase chromosomes lined up in descending order of their size (Figure 9. Karyotypes are prepared from actively dividing cells, such as white blood cells, bonemarrow cells, or cells from meristematic tissues of plants. After treatment with a chemical (such as colchicine) that prevents them from entering anaphase, the cells are chemically preserved, spread on a microscope slide, stained, and photographed. The photograph is then enlarged, and the individual chromosomes are cut out and arranged in a karyotype. For human chromosomes, karyotypes are often routinely prepared by automated machines, which scan a slide with a video camera attached to a microscope, looking for chromosome spreads. A karyotype for a male is shown here; a karyotype for a female would have two X chromosomes. Types of Chromosome Mutations Chromosome mutations can be grouped into three basic categories: chromosome rearrangements, aneuploids, and polyploids (Figure 9. Chromosome rearrangements alter the structure of chromosomes; for example, a piece of a chromosome might be duplicated, deleted, or inverted. In aneuploidy, the number of chromosomes is altered: one or more individual chromosomes are added or deleted. Some organisms (such as yeast) possess a single chromosome set (1n) for most of their life cycles and are referred to as haploid, whereas others possess two chromosome sets and are referred to as diploid (2n). A polyploid is any organism that has more than two sets of chromosomes (3n, 4n, 5n, or more). C D E takes a picture of the chromosomes, the image is digitized, and the chromosomes are sorted and arranged electronically by a computer. Preparation and staining techniques help to distinguish among chromosomes of similar size and shape. Duplications, trisomy, and autotriploids are examples of each category of mutation. G A B C D Rearranged chromosome (d) Translocation A D E F G M N O P Q R S B C D E F G (b) Deletion A B C In a chromosome deletion, a segment of the chromosome is deleted. M N O P E F S In a translocation, a segment of a chromosome moves from one chromosome to a nonhomologous chromosome (shown here) or to another place on the same chromosome (not shown). The four basic types of rearrangements are duplications, deletions, inversions, and translocations (Figure 9. The pairing and synapsis of homologous regions require that one or (a) Normal chromosome A B C D E F G Duplications A chromosome duplication is a mutation in which part of the chromosome has been doubled (see Figure 9. This type of duplication, in which the duplicated region is immediately adjacent to the original segment, is called a tandem duplication. If the duplicated segment is located some distance from the original segment, either on the same chromosome or on a different one, the chromosome rearrangement is called a displaced duplication. F E F G Alignment in prophase I of meiosis (b) A B C D E F G A B C D E F G E F Effects of chromosome duplications An individual homozygous for a duplication carries the duplication on both homologous chromosomes, and an individual heterozygous for a duplication has one normal chromosome and one chromosome with the duplication. Chromosome Variation 243 both chromosomes loop and twist so that these regions are able to line up (Figure 9. The appearance of this characteristic loop structure in meiosis is one way to detect duplications. Among fruit flies (Drosophila melanogaster), for example, a fly having a Bar mutation has a reduced number of facets in the eye, making the eye smaller and bar shaped instead of oval (Figure 9. The Bar mutation results from a small duplication on the X chromosome that is inherited as an incompletely dominant, X-linked trait: heterozygous female flies have somewhat smaller eyes (the number of facets is reduced; see Figure 9. Occasionally, a fly carries three copies of the Bar duplication on its X chromosome; for flies carrying such mutations, which are termed double Bar, the number of facets is extremely reduced (see Figure 9. The Bar mutation arises from unequal crossing over, a duplication-generating process (Figure 9. After all, gene sequences are not altered by duplications, and no genetic information is missing; the only change is the presence of additional copies of normal sequences. The answer to this question is not well understood, but the effects are most likely due to imbalances in the amounts of gene products (abnormal gene dosage). The amount of a particular protein synthesized by a cell is often directly related to the number of copies of its corresponding gene: an individual organism with three functional copies of a gene often produces 1. Because developmental processes require the interaction of many proteins, they often depend critically on proper gene dosage. If the amount of one protein increases while the amounts of others remain constant, problems can result (Figure 9. Although duplications can have severe consequences when the precise balance of a gene product is critical to cell function, duplications have arisen frequently throughout the evolution of many eukaryotic organisms and are a source of new genes that may provide novel functions. For example, humans have a series of genes that encode different globin chains, some of which function as an oxygen carrier during adult stages and others that function during embryonic and fetal development. All of these globin genes arose from an original ancestral gene that underwent a Wild-type chromosomes Chromosomes do not align properly, resulting in unequal crossing over. Bar chromosomes Unequal crossing over between chromosomes containing two copies of Bar. A B C Wild-type chromosome Gene expression Interaction of gene products 2 Development may be affected by the relative amounts of gene products. These techniques reveal that about 4% of the human genome consists of segmental duplications. In individuals heterozygous for a chromosome duplication, the duplicated region of the chromosome loops out when homologous chromosomes pair in prophase I of meiosis. Duplications often have major effects on the phenotype, possibly by altering gene dosage. Embryo Abnormal development 5 If the amount of one product increases but amounts of other products remain the same, developmental problems often result. A second type of chromosome rearrangement is a chromosome deletion, the loss of a chromosome segment (see Figure 9. A large deletion can be easily detected because the chromosome is noticeably shortened. In individuals heterozygous for deletions, the normal chromosome must loop during the pairing of homologs in prophase I of meiosis (Figure 9. This looping out generates a structure that looks very much like that seen for individuals heterozygous for duplications. Segmental duplications the human genome consists of numerous duplicated sequences called segmental duplications, which are defined as duplications greater than 1000 bp in length.
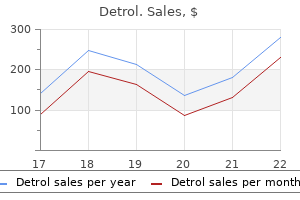
Generic 1mg detrol. Cardiology: Drug Induced Long QT syndrome (aLQTS).
References